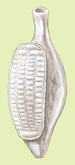

Aspidogaster conchicola
What do they look like?
The mature Aspidogaster conchicola is approximately 2.5 to 3.0 mm in length and 1.0 mm wide, and somewhat resembles a miniature conch. A single large ventral sucker, known as an opisthaptor, takes up most of the surface area of its underbelly. The opisthaptor is divided into adhesive depressions (termed loculi) formed by muscular septa, which are useful in classification. Aspidogaster conchicola has 64-66 loculi, arranged in four longitudinal rows. An exterior longitudinal septum, which is a horizontal flap of muscle, divides the body anteriorly. The function of the septum is unknown.
A wide buccal funnel (mouth) has its opening at the anterior end of a flexible neck-like process. Pharyngeal muscles move food entering the buccal funnel into the intestine. Unlike the related Digenean worms, aspidobothreans have a simple, unbranched digestive tract that ends in the cecum, a digestive sac surrounded by muscle. Wastes are removed by flame cell protonephridia, which channel material through excretory ducts into an excretory bladder, eventually exiting the body through a posteriorly located excretory pore.
An interesting feature of A. conchicola is its complex nervous system, a feature more characteristic of free-living rather than parasitic worms. Aspidogaster conchicola possesses an anterior cerebral commissure, an intricate organization of nerves. This presumably serves to coordinate the peripheral nerves, which are arranged in a ladder-type system. Numerous types of sensory receptors have been observed, particularly concentrated around the buccal funnel and opisthaptor, indicating that there is a good degree of neural coordination of locomotive and digestive action. (Bailey and Tompkin, 1971; Halton and Lyness, 1971; Hathaway, 1971; Roberts and Janovy Jr., 2000)
- Other Physical Features
- ectothermic
- heterothermic
- bilateral symmetry
-
- Average length
- 2.5 to 3 mm
- in
Where do they live?
Aspidogaster conchicola is a parasite of freshwater bivalves, with a wide distribution in North America, Africa, and Europe. This small fluke has a low degree of host specificity and has been found in a wide variety of other hosts, including snails, fish, and turtles. Inside the typical molluscan host, the parasite will usually take residence within the pericardial cavity. (Huehner and Etges, 1977; Rohde, 1972)
What kind of habitat do they need?
Not being well adapted to any single host species, A. conchicola will readily develop in a wide variety of freshwater mollusk, snail, fish, or turtle families. In laboratory settings A. conchicola was able to survive in water for two weeks without a host, and in a saline solution for up to five weeks. This suggests that Aspidogaster is an archaic species, not too far removed from a free-living ancestor. (Rohde, 1972; Rohde, 1998)
- These animals are found in the following types of habitat
- freshwater
- Aquatic Biomes
- lakes and ponds
- rivers and streams
How do they reproduce?
Aspidogaster conchicola usually develops within mollusks, but is a facultative parasite of vertebrates, should their mollusk host be eaten. Their life cycle is still direct, requiring only one host for maturation. Infection occurs through intake of the egg, containing a fully developed larva, by the mollusk's siphon. Upon entering the host, the cotylocidium (larvae) will hatch and immediately begin maturation without further migration. At hatching, the larvae are 13 to 17 micrometers long. Since the cotylocidium does not seek the host as in other trematodes, it is unciliated. Instead, it has a simple posterior sucker lacking loculi. There have been no documented cases of Aspidogaster directly infecting a vertebrate host as a free egg or larva, while they have been found in the intestines of fish, so it is assumed that vertebrates are infected by ingestion of parasitized mollusks.
Aspidobothreans are able to either self-fertilize or mate with another individual. (Bakker and Davids, 1973; Huehner and Etges, 1977; Rohde, 1972; Rohde, 1998)
- Key Reproductive Features
- simultaneous hermaphrodite
- sexual
- fertilization
- ovoviviparous
- oviparous
- Parental Investment
- no parental involvement
How do they behave?
The large ventral opisthaptor appears to play a major role in Aspidogaster conchicola feeding behavior. Muscular action mechanically disrupts host tissues while marginal organs within the sucker release secretions for extracorporeal digestion. The predigested material is then sucked in through the buccal funnel.
Aspidogaster infections appear to be most prevalent and intense during the colder months, from November through April. (Bailey and Tompkin, 1971; Halton and Lyness, 1971; Huehner, 1987; Huehner, et al., 1989)
- Key Behaviors
- parasite
What do they eat?
The primary hosts for Aspidogaster conchicola are freshwater mollusks, usually mussels. However, as a facultative parasite of vertebrates, it is not restricted to mussels, and will also enter into and develop within freshwater snails, fishes, and turtles while feeding on epithelial tissue. (Rohde, 1972)
How do they interact with us?
The aspidobothreans do not hold any economic or medical significance for humans. However, they have several archaic characteristics which suggest that they are an ancient group, and so continue to be studied because they seem to represent a link between parasitic and free-living organisms. Since Aspidogaster has not developed any close associations with particular host species, it has been suggested that it is an archaic species of trematode, similar to the hypothetical ancestor from which the related digenean trematodes have evolved their complex life cycles exploiting multiple hosts. (Rohde, 1972)
Aspidogaster conchicola infects the zebra mussel, Dreissena polymorpha, which is an invasive species in the Great Lakes in the United States. (Laruelle and Mollogy, 1996)
Are they endangered?
-
- IUCN Red List
- No special status
-
- US Federal List
- No special status
-
- CITES
- No special status
Contributors
Renee Sherman Mulcrone (editor).
Raymond Pahk (author), University of Michigan-Ann Arbor, Solomon David (editor), University of Michigan-Ann Arbor.
References
University of Manitoba. 2000. "Cotylogaster occidentalis" (On-line). WELCOME TO PARASITOLOGY 22.346. Accessed November 14, 2003 at http://www.umanitoba.ca/faculties/science/zoology/faculty/dick/z346/aspidohome.html.
Bailey, H., S. Tompkin. 1971. Ultrastructure of the integument of Aspidogaster conchicola. Journal of Parasitology, 57: 848-854.
Bakker, K., C. Davids. 1973. Notes on the life history of Aspidogaster conchicola Baer (Trematoda; Aspidogastridae). Journal of Helminthology, 47: 269-276.
Halton, D., R. Lyness. 1971. Ultrastructure of the tegument and associated structures of Aspidogaster conchicola (Trematoda; Aspidogastrea). Journal of Parasitology, 57: 1198-1210.
Hathaway, R. 1971. The fine structure of the trematode Aspidogaster conchicola von Baer, 1827. Dissertation Abstracts International, 31B: 7687.
Huehner, M. 1987. Aspidogastrid and digenetic trematode single an double infections in the gastropod Elimia livescens from the Upper Cuyahoga River, Ohio USA. Proceedings of the Helminthological Society of Washington, 54 (2): 200-203.
Huehner, M., F. Etges. 1977. The life cycle and development of Aspidogaster conchicola in the snails, Viviparus malleatus and Goniobasis livescens. Journal of Parasitology, 63: 669-674.
Huehner, M., K. Hannan, M. Garvin. 1989. Feeding habits and marginal organ histochemistry of Aspidogaster conchicola (Trematoda; Aspidogastrea). Journal of Parasitology, 75 (6): 848-852.
Laruelle, F., D. Mollogy. 1996. A Guide to Identifying the Endosymbionts of Dreissena polymorpha. The 6th International Zebra Mussel and Other Aquatic Nuisance Species Conference, Dearborn, Michigan. Accessed November 14, 2003 at http://sgnis.org/publicat/96Laruel.htm.
Roberts, L., J. Janovy Jr.. 2000. Foundations of Parasitology, 6th ed. New York: McGraw Hill.
Rohde, K. 1998. "Aspidogastrea" (On-line). Accessed 11/14/03 at http://ag.arizona.edu/ENTO/tree/eukaryotes/animals/platyhelminthes/aspidogastrea/aspidogastrea.html.
Rohde, K. 1972. The Aspidogastrea, especially Multicotyle purvisi Dawes, 1941. Advances in Parasitology, 10: 77-151.


BioKIDS home | Questions? | Animal Diversity Web | Cybertracker Tools
Pahk, R. 2003. "Aspidogaster conchicola" (On-line), Animal Diversity Web. Accessed April 23, 2024 at http://www.biokids.umich.edu/accounts/Aspidogaster_conchicola/BioKIDS is sponsored in part by the Interagency Education Research Initiative.
It is a partnership of the University of Michigan School of Education, University of Michigan Museum of Zoology, and the Detroit Public Schools. This material is based upon work supported by the National Science Foundation under Grant DRL-0628151.
Copyright © 2002-2024, The Regents of the University of Michigan. All rights reserved.